In the first article "Origin of Life: A Critique of Early Stage Chemical Evolution Theories,", January, 1976, of this series on origin of life theories, following the discussion of problems involved in a naturalistic origin of relatively simple organic compounds, the problem of the origin of large molecules ("macromolecules"), such as the proteins, DNA, and RNA, was introduced. It was pointed out that one of the insuperable barriers to the accumulation of significant quantities of these very complex molecules (even assuming that the ocean was populated with huge quantities of the necessary chemicals) was the fact that energy is required to form the chemical bonds between the units in these long-chain compounds.
 |
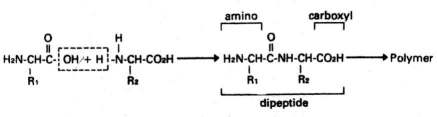
As a consequence, there is, practically speaking, no tendency for these compounds to form, but, on the other hand, they very readily tend to fall apart or disintegrate. What happens naturally and spontaneously, then, is that proteins break up into their constituent amino acids, and DNA and RNA tend to break up into fragments, and eventually into their constituent groups -- a sugar, phosphoric acid, and purines and pyrimidines. If proteins, DNA, RNA, and other complex macromolecules arose on the hypothetical primitive earth by naturalistic processes, some mechanism would have had to exist to drive this process in the direction opposite to that which it tends to go. This mechanism would have had to be very efficient, since many billions of tons each of many different kinds of proteins, DNA, and RNA would have to be produced to provide enough of these vital compounds for the origin of life in an ocean containing somewhere between 35 and 350 million cubic miles of water.
Fox's Thermal Model
The suggestion that has gained more attention than all others is the idea of Sidney Fox. Fox has published papers on various aspects of his thermal theory in numerous scientific journals and in many books, a few of which are listed in the bibliography of this paper.1-5 An outline of Fox's theory can be found in practically every modern high school and college text on biology, evolution, and related subjects. Recently a review volume was published in honor of his 60th birthday.6 And yet if any thing in science is certain, it can be said that however life arose on this planet, it did not arise according to the scheme suggested by Fox. One could not be judged to be too unkind or critical if he were to label Fox's suggestion as pseudoscience.
Fox uses intense heat as the driving mechanism in his model. In the laboratory demonstration of Fox's origin of life scheme, a particular mixture of pure, dry amino acids are heated at about 175° C (water boils at 100° C) for a limited time (usually about six hours). Intense heating is then ceased, and the product is stirred with hot water, and insoluble material is removed by filtration. When the aqueous solution cools, a product precipitates in the form of microscopic globules, which Fox calls proteinoid microspheres. Analysis of this material shows that it consists of polymers, or chains, of amino acids, although of shorter lengths than are usually found in proteins. Some of these globules resemble coccoid bacteria, and others bulge and superficially appear to be budding similar to certain microorganisms.
Fox claims that his proteinoid microspheres constitute protocells (that is, they are almost, but not quite, true cells), and were a vital link between the primordial chemical environment and true living cells. He claims that the amino acids in these polymers are not randomly arranged as would be expected, but that a few highly homogeneous (having identical chemical structure) protein-like molecules are obtained with their amino acids arranged in a precisely ordered sequence. He further claims that these compounds possess detectable catalytic or enzyme-like properties. Finally, Fox claims that these microspheres multiply by division somewhat in the manner of true cells.
 |
When asked where on the primordial earth a locale could be found where amino acids might have been heated at about 175° C, Fox suggests that such a locale would have been found on the edges of volcanoes. When it was pointed out that heating at that high a temperature (not much reaction occurs at temperatures much below 175° C) would cause complete destruction of the products if heating continues much beyond six hours, Fox suggests that rain might occur just at the right time to wash away the products.
Fox's scheme would require such a unique series of events and conditions, the probability of which would be so vanishingly small that it could be equated to zero. These are the following:
1. Heating at a high temperature for a limited amount of time.
Fox's suggestion that the combination of the edges of volcanoes with rain at just the right time would suffice to produce billions of tons of these polymers has been severely criticized even by numerous evolutionists.7 Miller and Orgel point out that when lava solidifies, the surface of the lava is hardly warmer than air temperature. In discussing this feature of Fox's model they say, "Another way of examining this problem is by asking whether there are places on the earth today with appropriate temperatures where we could drop, say, 10 grams of a mixture of amino acids, and obtain a significant yield of polypeptides… We cannot think of a single such place."8 Even if there were such places, they would be so limited in extent, and the timing of the rain would be so restrictive (not much less nor much more than six hours from the time heating begins), that the rate of production would be very much less than the rate of destruction by hydrolysis and other degradative reactions once the products were washed into the ocean or other bodies of water.
2. Fox's reaction mixture consists solely (as far as organic material is concerned) of pure amino acids.
Where on earth could a mixture of pure amino acids be found? Only in the laboratory of a twentieth-century scientist! According to the chemical evolutionary scheme to which Fox and every other origin of life theorist subscribes, however, a great variety of organic chemical compounds, numbering in the thousands and most likely many tens of thousands, would have been produced on the primordial earth. The probability of a mixture of pure amino acids accumulating anywhere, assuming that they were being produced, would be absolute zero. Any amino acids produced would be admixed with sugars, aldehydes, ketones, carboxylic acids, amines, purines, pyrimidines, and other organic chemicals. Heating amino acids at almost any temperature with a mixture of such chemicals would be certain to result in complete destruction of the amino acids. Beyond question, no polypeptides or proteinoids would be produced. This factor alone completely eliminates Fox's scheme from any rational discussion.
3. A totally improbable ratio of amino acids is required.
If random proportions of amino acids are heated, no product is obtained. A very high proportion of one of the acidic amino acids, aspartic and glutamic acids, or of the basic amino acid, lysine, is required. Generally, about one part of one of the acidic amino acids, or one part of lysine, a basic amino acid, is heated with two parts of all the remaining amino acids combined. Under no naturally occurring conditions would any such ratio of amino acids ever exist. In all origin of life laboratory experiments, the amino acids produced in highest ratios are glycine and alanine, the simplest in structure and therefore the most stable of all the amino acids. Aspartic and glutamic acids are generally produced, but in small proportions. Detectable quantities of lysine are rarely, if ever, produced. Again, Fox's scheme is completely out of touch with reality.
4. Serine and threonine are mainly destroyed.
Two of the most commonly occurring amino acids in proteins consist of serine and threonine. Yet they undergo severe destruction during the heating process required in Fox's scheme. The resultant product thus contains only minor amounts of serine and threonine in contrast to naturally occurring proteins.
5. The claim that the products consist of a few relatively homogeneous polypeptides ("proteinoids") with amino acids arranged in a highly ordered sequence is patently absurd.
If a monkey were allowed to type away on a typewriter, the sequence in the string of letters produced on the paper would be completely random. The result would be nonsense. So it is with polymers produced from amino acids, nucleotides, or sugars according to ordinary chemical and physical processes. Chemistry and physics, just like monkeys, are dumb things, and have no ability to arrange subunits in any particular order. Probability considerations based on relative reactivities of functional groups and activation energies require the production of random structures or sequences in any polymerizations involving mixtures of amino acids, nucleotides, or sugars. It has been demonstrated that, in fact, polymerization of sugars9 and of nucleotides10 leads to random sequences.
Fox's claim that his product consists of relatively large quantities of a few polypeptides (polymers of amino acids are called polypeptides when the chains are shorter than proteins), each with the amino acids arranged in a highly specific sequence, rather than an enormous number of polypeptides with random structures, is based upon entirely inadequate separation techniques and analyses. There is no valid evidence whatever to show whether or not the amino acids in Fox's products are ordered. In fact, some of his fellow origin of life theorists accuse Fox of deception in this respect. Thus, Miller and Orgel, concerning Fox's claim that his product consists of nonrandom polypeptides, say "Thus, the degree of nonrandomness in thermal polypeptides so far demonstrated is minute compared with the non-randomness in proteins. It is deceptive, then, to suggest that thermal polypeptides are similar to proteins in their nonrandomness."11
Beyond the above considerations, there is additional compelling evidence that Fox's product must consist of random structures. The high temperature required for the reaction nearly completely racemizes the amino acids. All but one of the amino acids found in proteins (glycine is the exception) may exist in at least two forms, forms in which the arrangement in space of the atoms differ. These forms are designated as the D- and L- forms (sometimes called "right-" and "left-handed"). They bear the same relationship to each other that a right hand bears to a left hand; each is a mirror-image of the other but not superimposable. Chemically and physically they exhibit identical properties except that solutions of the two forms rotate plane-polarized light in opposite directions. Biologically the difference is enormous, however. All naturally occurring proteins contain exclusively the L-, or "left-handed," form. The replacement of a single amino acid in a protein with its D-form completely destroys all biological activity.
 |
Racemization is the process which converts D-amino acids to a mixture of the D- and L-forms, or L-amino acids to a mixture of the D- and L-forms. When an amino acid is completely racemized it consists of equal quantities of the D- and L-forms. All amino acids tend to racemize under natural conditions, the rate of racemization depending on the particular amino acid and environmental conditions. The brutal treatment of heating amino acids several hours at 175° C, as mentioned above, extensively racemizes the amino acids, changing the amino acids from L-forms to a mixture of L- and D-forms.
Since the D- and L-forms of amino acids have identical chemical properties, the probability of the D-form being incorporated at any point in the chain is equal to the probability of the incorporation of the L-form. There would be no way then, chemically, of specifying which form would be incorporated at any particular point. The sequence of the first two amino acids in the chain might thus be L-L, D-D, D-L, or L-D. Each would have equal probability. The sequence of the first three amino acids, whatever the particular amino acids, might be L-L-L, L-L-D, L-D-L, L-D-D, D-D-D, D-D-L, D-L-D, or D-L-L. Thus, it can be seen that even if the sequence of the first three amino acids were the same (such as, for example, arginine-valine-threonine), eight different structures can be obtained, differences which would exert enormous influence biologically. In fact, based on known bio-chemistry, only the L-L-L form could have had any potential significance.
It is thus impossible for Fox's product to consist of specific structures. A particular sequence of ten amino acids but consisting of mixtures of the D- and L-forms would yield a thousand different structures (210) and a particular sequence of 100 amino acids existing in D- and L-forms would yield 10 billion times 10 billion times 10 billion different structures (2100 or approximately 1030). It is apparent that Fox's claim for a high degree of homogeneity, or non-randomness, in his product is indeed absurd.
6. Catalytic, or enzymic, properties claimed for the product are barely detectable and unrelated to present enzymes.
The catalytic properties of enzymes found in present-day organisms are due to the precise sequence of the L-amino acids in these proteins. Fox's product consists of random sequences of these amino acids. Any enhancement of the catalytic activity of the free amino acids themselves by this polymerization would be no more than that conveyed by the incorporation of these amino acids into random polymers or nonspecific chemical structures. Furthermore, these polymers consist of mixtures of D- and L-amino acids. As mentioned earlier, the substitution of only one L-amino acid by its D-form in an enzyme (which may consist of several hundred amino acids) completely demolishes, for all practical purposes, its biological, that is, its catalytic, ability (residual activity, if any, is reduced below a detectable quantity). Further discussion of this point may be found in the monograph by Gish on the origin of life.12 It is probable that if Fox had swept up the dust on the floor of the university administration building and thrown it into his test mixture, it would have had as much activity as his proteinoid.
7. The proteinoid microspheres are unstable and are easily destroyed.
Fox claims a rather high degree of stability for his proteinoid microspheres, yet he, himself, reveals that microspheres contained in aqueous suspension between microscope slides can be easily redissolved by merely warming the slides.13 Stable, indeed! Furthermore, dilution of an aqueous suspension by adding water also dissolves the microspheres.
8. Division of the microspheres is due to simple physicochemical phenomena and have no relation to cell division by living organisms.
Cell division in even the simplest organisms requires an incredibly complex process and machinery, involving duplication of each unit of the cell with extremely high fidelity. On the other hand, the division reported for Fox's microspheres is a simple physicochemical phenomenon, like the separation of a soap bubble into two bubbles. It has no greater significance. As material precipitates from solution in the form of globules, and as the quantity that has collected in any particular globule exceeds a certain amount, physicochemical forces may cause the globule to split into two globules. No reproduction, no replication of any kind, however, takes place. The material in the first globule would be randomly distributed between the two product globules.
This discussion of the Fox scheme for the origin of life, even though incomplete, has been relatively extensive. This is believed desirable, however, because of the tremendous promotion (and naive acceptance) of Fox's theories in high school and college texts and in scientific circles as well. Fox's success confirms the bias and unscientific attitudes that dominate the educational and scientific establishments in relation to the question of origins. Anything that incorporates evolutionary philosophy is acceptable, no matter how unscientific.
Other Models
Other suggestions have been offered (good but concise reviews of these may be found in the paper by Horowitz and Hubbard7b and the book by Miller and Orgel7a). Those that involve reactions in aqueous solution (and thus in the oceans, lakes, and all other aqueous environments) can be effectively eliminated because the high energy reagents required to provide the energy to form the chemical bonds between the amino acids, nucleotides, etc., would be rapidly destroyed by water. These reagents are effective in laboratory syntheses because the reagents are prepared in non-aqueous solvents under anhydrous conditions, and the reactions in which these reagents are used are generally carried out under similar conditions. There is no possibility that these reagents could form on the primitive earth, however.
Other suggestions utilizing elevated temperatures in a dry environment, in addition to the suggestion of Fox, have been offered.14 Orgel and his collaborators have published a series of papers, for example, on the thermal synthesis in a dry environment of nucleotides and of polymers of nucleotides,15 but Orgel, himself, admits that these experiments have no relevance to the origin of life. After discussing the possibilities of such reactions occurring under primitive earth conditions, Miller and Orgel state, "However, we doubt that very extensive polymerization of nucleotides could have occurred in this way, or that 'biological' polymerization could have taken place except in an aqueous environment."16
Miller and Orgel have thus stated their conviction that polymerizations that gave rise to proteins, DNA, RNA, and other biological molecules ("‘biological’ polymerizations") must have occurred in an aqueous environment. But as stated above, this would have been impossible because the high energy compounds needed to drive these polymerization reactions could not have formed or existed in an aqueous environment.
In the concluding paragraph to their chapter on polymerizations, Miller and Orgel state, "This chapter has probably been confusing to the reader. We believe this is because of the very limited progress that has been made in the study of prebiotic condensation reactions."17 This lack of success has resulted from the extreme difficulties in attempting to imagine how such processes could have occurred under natural conditions. Some might suppose, on the other hand, that limited progress has been made mainly because comparatively little research has yet been done on the origin of life. In that limited amount of research, however, enough work has been done to test all principles involved. Further work will not alter the principles of thermodynamics, chemical kinetics, or other basic principles involved. These stand as barriers to a naturalistic origin of biologically active molecules.
This series on origin-of-life theories will be concluded in a future issue.
REFERENCES
1 S. W. Fox, Ed., The Origins of Prebiological Systems and of Their Molecular Matrices, Academic Press, New York, 1965.
2 S. W. Fox and K. Dose, Molecular Evolution and the Origin of Life, Freeman Pub. Co., San Francisco, 1972.
3 S. W. Fox, "Self-ordered Polymers and Propagative Cell-Like Systems," Naturwissenschaften, Vol. 56, pp. 1-9 (1969), in English.
4 S. W. Fox, K. Harada, G. Krampitz, and G. Mueller, "Chemical Origins of Cells," Chemical and Engineering News, June 22, 1970, pp. 80-94.
5 S. W. Fox, "The Protein Theory of the Origin of Life," American Biology Teacher, Vol. 36, pp. 161-172 (1974).
6 D. L. Rohlfing and A. I. Oparin Eds., Molecular Evolution: Prebiological and Biological, Plenum Press, New York, 1972.
7 (a) S. L. Miller and L. E. Orgel, The Origins of Life on the Earth, Prentice-Hall, Englewood Cliffs, N.J., 1973, p. 145; (b) N. H. Horowitz and J. S. Hubbard, "The Origin of Life," Annual Review of Genetics, Vol. 8, p. 399 (1974); (c) C. Sagan, in Reference 1, p. 374;
d) J. R. Vallentyne, in Reference 1, p. 379.
8 Reference 7a, p. 145.
9 P. T. Mora, in Reference 1, p. 287.
10 G. Schramm, in Reference 1, p. 299.
11 Reference 7a, footnote on p. 144.
12 D. T. Gish, Speculations and Experiments Related to Theories on the Origin of Life, Creation-Life Publishers, San Diego, 1972.
13 S. W. Fox, "A Theory of Macromolecular and Cellular Origins," Nature, Vol. 205, p. 336 (1965).
14 See for example Reference 7a, pp. 141-148, and Reference 7b, pp. 399-400.
15 For a brief review of this work, see Reference 7b, p. 400.
16 Reference 7a, p. 142.
17 Reference 7a, p. 148.*Dr. Gish is Vice President of ICR.



