COMPLEX LIFE CYCLES IN HETEROPHYID TREMATODES: STRUCTURAL AND DEVELOPMENTAL DESIGN IN THE ASCOCOTYLE COMPLEX OF SPECIES
MARK H. ARMITAGE, M.S.
ICR Graduate School, 10946 Woodside Ave. N., Santee, CA 92071
Voice: (619) 448-0900 Fax: (619) 448-3469
Voice: (619) 448-0900 Fax: (619) 448-3469
Presented at the Fourth International Conference on Creationism
Pittsburgh, PA, August 3-8, 1998
Copyright 1998 by Creation Science Fellowship, Inc.
Pittsburgh, PA USA - All Rights Reserved
Pittsburgh, PA, August 3-8, 1998
Copyright 1998 by Creation Science Fellowship, Inc.
Pittsburgh, PA USA - All Rights Reserved
KEYWORDS
Trematode, obligate parasite, intermediate host, cercaria, metacercaria, life cycles, design, chance.
ABSTRACT
Microscopic heterophyid trematode parasitic worms of the Genus
Ascocotyle infect certain amnicolid and hydrobiid snails and certain
cyprinodont and poeciliid estuarine fishes as first and second
intermediate hosts. Adult trematode worms are found to mature
in the intestines of particular definitive hosts, most often piscivorous
birds, but also certain mammals. A survey of these parasites,
harvested from fish hearts and gills collected in Mississippi,
Texas and California, shows that they are obligated to complex
life cycles requiring at least three disparate and different hosts
to achieve fecundity. Methods of infection, host infection site
and host specificity are often unique to each different species
of these parasites. Additionally, Ascocotyle worms demonstrate
highly specialized structures such as HCl resistant cysts, HCl
sensitive penetration glands and sensory organs which may serve
to guide them to the specific infection site. These heteroecious
life cycles and specialized structures are shown to be too complex
to have developed by chance, therefore, evolutionary mechanisms
appear insufficient to explain them. A creationist design argument
for the presence of such parasites is promulgated.
INTRODUCTION
Arthur Looss [1] erected the genus Ascocotyle with A. coleostomum
(Looss, 1896) as the genotype for the Ascocotyle complex. Excluding
synonyms, or identical organisms with differing names, there are
at present over 30 named species comprising the Ascocotyle complex
as defined per Travassos [2], Stunkard and Uzmann [3], Burton
[4], Sogandares-Bernal and Bridgman [5], and Sogandares-Bernal
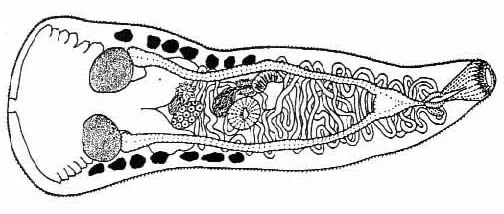
and Lumsden [6]. The adult worms (Fig #1) are non-pathogenic, intestinal parasites of piscivorous,
or fish-eating, birds and mammals [7].
{kind=link}
It is in the intestines of these definitive hosts that the parasite
matures and produces eggs which are passed with feces into the
marsh or estuary [8]. Because these organisms are hermaphroditic,
or self-fertilizing, one parasite can populate a marsh with eggs
[9]. Each fertilized egg has been shown to be able to produce
a redia capable of growing 6000 swimming larvae over a one year
period [10]. The first intermediate hosts are amnicolid and hydrobiid
snails, which take up the eggs while feeding over the bottom surface
of the estuary [11]. Infection within the particular snail type
specifically depends upon the particular species of Ascocotyle
present, and cross infection has been shown not to occur in many
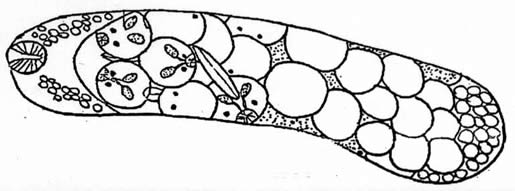
experimental trials [12, 13, 14, 15]. The eggs develop into a
redia or brood stage (Fig
#2), and often travel from the the snail digestive gland to
the gonads or hepatopancreas where further development ensues.
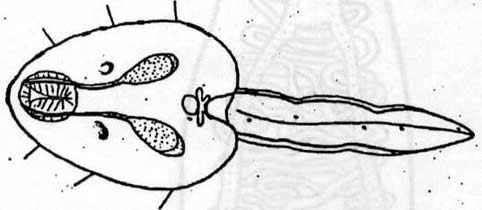
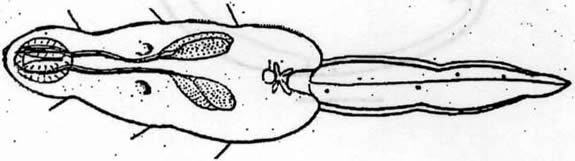
The redia grows in length and bears up to 50 cercariae in various
stages of development. When mature, these cercariae (Fig
#3, Fig #4)
leave the redia through a birth pore, mostly during daylight hours
(but often at dawn), and swim out of the snail towards illuminated
areas of the water [16, 17]. The second intermediate hosts are
centrarchiid, cyprinodontid, mugilid, and poeciliid fishes, and,
in at least one instance, anuran tadpoles [18]. Cercariae swim
near these fish and are taken up in the respiratory current. Some
Ascocotyle will, at this point, attach to and penetrate the gill
filaments of the fish, dropping their tails before or soon after
penetration [19, 20, 21]. Some species will immediately encyst
in the gill, by the production of a multi-layered hyaline, collagen-like
cyst [22, 23], while others will search for and enter the efferent
blood vessels supplying blood to the gills [24, 25]. The metacercariae
will then travel within the blood system, either against or with
blood flow, to the heart, liver, brain and other organs, where
cysts will be produced and further development is arrested [ibid].
Other species of cercariae do not attach to the fish gills, but
rather, are swallowed by the fish. They produce cysts in the stomach
and intestine or penetrate the intestine and encyst in the liver
and mesentary [26, 27].
{kind=link}
{kind=link}
{kind=link}
As mentioned previously, host infection and even organ infection
site is Ascocotyle species characteristic [28, 29, 30, 31]. It
is at this point in the life cycle, that predation upon the infected
fish by herons, egrets, or raccoons and other mammals, must occur
in order that the definitive host can become infected and the
worm can sexually mature. Metacercarial cysts, which are HCl resistant,
successfully pass the definitive host stomach and dissolve only
within the intestine, freeing the worm to fertilize and eject
eggs. These life cycles are typically completed in brackish water
estuarine marsh habitats, but some second intermediate hosts have
been shown to swim out to sea [32]. In this paper, specific, complex
life cycles and morphological structures of Ascocotyle leighi,
A. pachycystis and A. diminuta, collected from fish hearts in
Mississippi, Texas and California are evaluated in the light of
the creation/evolution paradigms.
MATERIALS AND METHODS
Sheepshead minnow, (Cyprinodon
variegatus), Sailfin Molly, (Poecilia latipinna), Mosquitofish
(Gambusia affinis) (Fig
#5) and Killifish (Fundulus parvipinnis) (Fig
#6) specimens were collected by seine, dip net and trap method
from Pine Island, Mississippi, South Padre Island, Texas, and
Newport Beach and Point Mugu, California, and of 60 fish collected,
only 7 were uninfected. The hearts, gills and livers of these
fish were harvested and examined under a dissecting microscope
for the presence of metacercarial cysts, indicating a possible
Ascocotyle infection. Some of the hearts and gills were fixed
in glutaraldehyde, embedded in plastics and thin-sectioned [33].
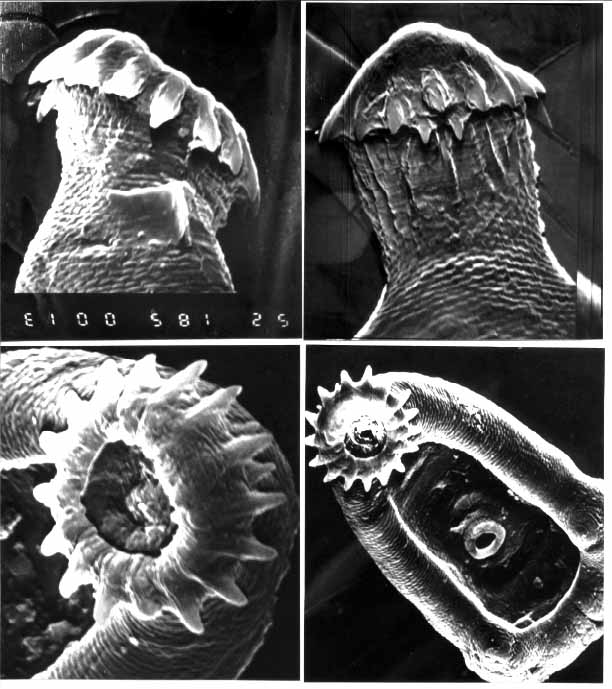
Some cysts were removed from the heart and gill tissue for mechanical
and enzymatic excystment and further study under scanning electron
microscopy [34], (Fig
#7). Worms which were enzymatically excysted only exited their
cysts under conditions of a 7.4 PH adjusted mixture of saline,
RPMI media and trypsin in an incubator at 370C for 3-5 hours.
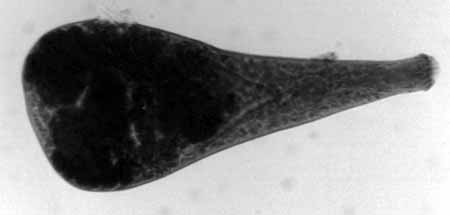
Using fine needle forceps, other cysts were forcibly popped open
releasing the live metacercaria for observation under high magnification
light microscopy. Whole mounts were made of some of these excysted
worms (Fig #8).
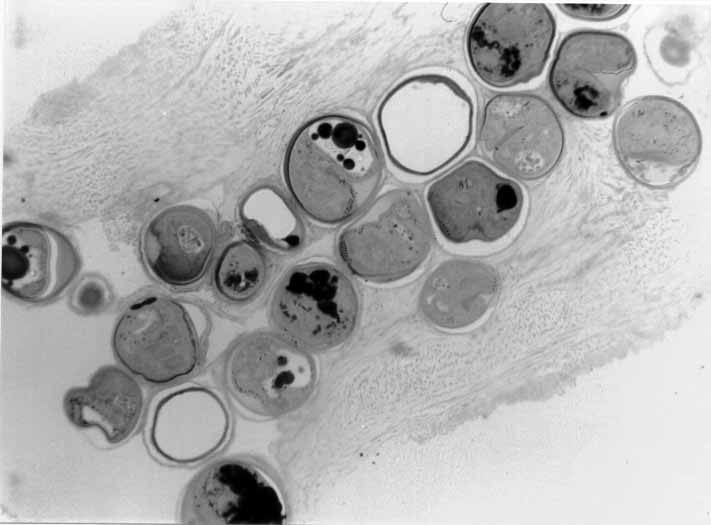
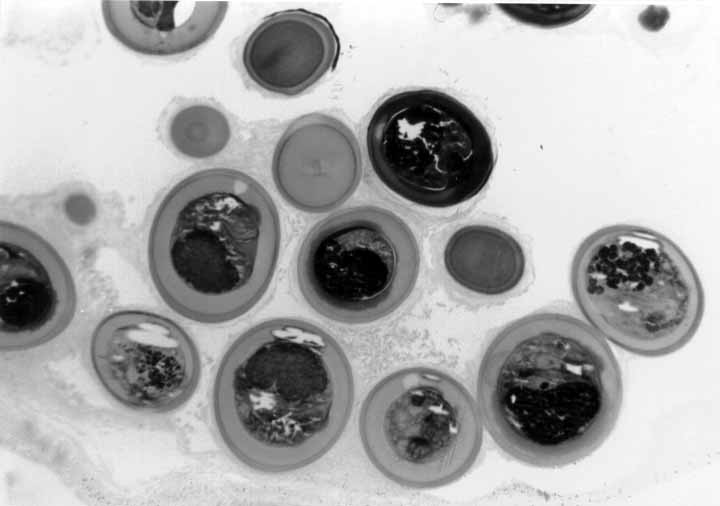
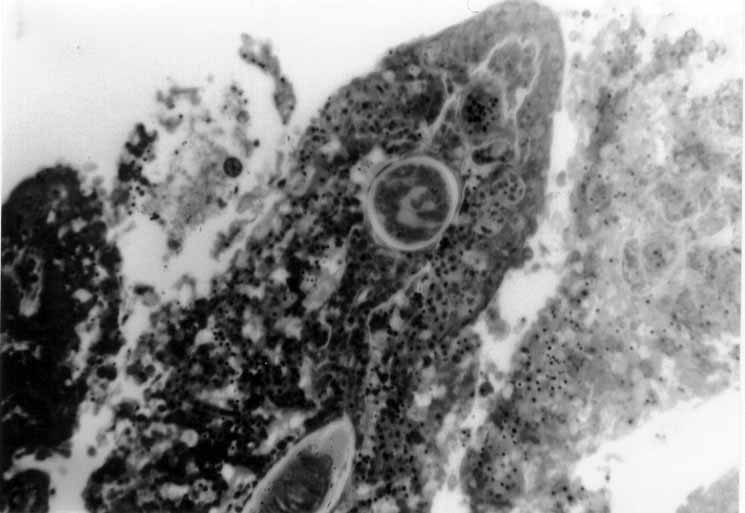
2-5 micron thick sections of fish hearts and gills were cut with
diamond and glass knives, stained with Methylene Blue and Azure
II and coverslipped (Fig
#9, Fig #10,
Fig #11).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Trematodes for SEM study were processed through osmium, thiocarbohydrazide
and a graded series of alcohols to absolute alcohol. Dehydration
by the critical point drying method was attempted resulting in
the loss of many specimens. Air dried worms were transferred by
hand under the dissecting microscope to SEM stubs and were sputter
coated for 4 minutes at 30mA deposition. Stubs were then transferred
to a Jeol JSM 35 Scanning Electron Microscope and were observed
and photographed at 100-2000X magnification at 15 and 25 KV.
DISCUSSION
These parasites are an evolutionary enigma for several reasons
and their presence raises more questions than are answered. The
evolutionary paradigm for r-strategists (or organisms which are
small, fast-growing and which have short, highly-populated generation
times), calls for them to employ a rapidly developing, independent
life cycle which allows them to exploit their environment quickly,
achieve sudden fecundity and bear the most possible offspring
with a minimum of exposure to survival hazards and expenditure
of energy. In the light of that definition, Ascocotyle breaks
all the rules. For one, Ascocotyle manifests a life cycle which
may take up to a year or much more to complete, if the second
intermediate host (bearing many such parasites) can escape predation
or death. As mentioned previously, many examples exist of infected
first and second intermediate hosts being kept alive in laboratory
aquaria for long periods of time with no ill-effect, showing that
the parasite is capable of surviving (or enduring) a lengthy hiatus
before it can pass on its genes. This type of life cycle is uncommon
for many microscopic organisms which typically bear offspring
quickly, and do not expend energy on many intermediate stages.
This may run counter to some thinking that evolution selects those
r-strategists which develop into fitter populations faster than
others in order to exploit the available environmental resources
[35]. Conversely, some authors feel that evolutionary selection
may run along a continuum from r-strategists to K-strategists
(slow-growing organisms with long generation times and few offspring),
where, "In the ecological void the optimal adaptive strategy
channels all possible resources into survival and production of
a few offspring of extremely high competitive ability." [36]
Clearly ascocotylids are not k-strategists in the sense that many
cercariae can be produced from one snail bearing a redia. By placing
most of those cercariae into one or a few hosts which may or may
not become predated upon, Ascocotyle certainly seems to minimize
its chances for success.
Secondly, Ascocotyle is anything but independent, being strictly
an obligate parasite. In the world of parasites, this feature
is not unique as many completely depend upon other organisms to
survive (many organisms would vanish tomorrow if the host population
upon which they live died out).
The enigma for a 'survival of the fittest' interpretation of
life cycles is that Ascocotyle, like most trematodes is obligated
to three hosts, and therefore its chances of reaching sexual maturity
are several times smaller than other, more independent organisms.
Again, this does not seem to square with the typical r-strategy
scenario within which ascocotylids should operate.
This parasite requires a snail as the first intermediate host,
but not just any snail will do. Some 160 types of snails inhabit
these estuaries [37], yet the rediae do not develop in most of
them. What is it about the "right" snail which causes
the egg to develop into a redia, and what happens to eggs which
are ingested by snails within which no development takes place?
It would seem best to develop an evolutionary plan which would
allow many types of intermediate hosts to serve as appropriate
vehicles for development.
It also appears, based on the life histories referenced herein
that the redia only develop in a certain part of the snail [38].
What mechanism guides the redia to the hepatopancreas or for that
matter the gonads for completion of that stage?
These parasites require a 2nd intermediate fish host and not
just any fish. As mentioned previously, laboratory studies have
shown that Ascocotyle is very selective about the fish hosts which
it infects.
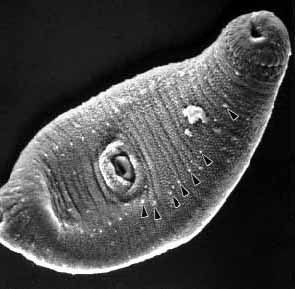
What is not clear is what mechanism guides these cercariae to
the proper fish for encystment. Do redial and cercarial spines
and sensory papillae around the oral sucker and along the ventral
tegument (Fig
#12) play a role in host/organ detection and emplacement?
When one would expect sibling species to be most alike, why do
some sibling species (like A. leighi and A. pachycystis) only
infect certain, but not the same fish in the same locality?
{kind=link}
In addition, ascocotylids are not only host specific but they
are organ specific within the 2nd intermediate host. Some metacercariae
will travel with blood flow within the 2nd intermediate host and
always end up in the same organ or site, others swim against the
efferent flow and encyst within the heart. What mechanism or sensory
apparatus indicates to the parasite which organ they are in or
how to get there? Since they are generally non pathogenic and
since a heavy parasite burden (even to the heart) has a minimal
impact upon the 2nd intermediate host [39] one wonders if these
parasites may confer some advantage to the host.
Finally, they require a 3rd host, which really begs the evolutionary
question. Concern over this is expressed by some authors, in an
attempt to supply an evolutionary explanation [40, 41]. Here the
definitive host must digest the fish, while the cyst must pass
this process unscathed. If the goal is to quickly survive and
reproduce, why tie survival to the (potentially lethal) digestive
process of a mammal or bird? It would seem to be "safer"
to infect, say the shell of a shellfish which may be discarded
by a raccoon or a bird after the meal is complete.
A. sexidigita and A. mcintoshi both appear to go the digestive
route [42, 43], not once, but twice; first within the fish and
then the bird. What allows this cercaria to resist digestion within
the fish stomach if it does not encyst within a protective capsule
before burrowing through the intestinal submucosa to the visceral
organs?
Cysts ingested by the definitive host also do not dissolve in
HCl which is found normally within the host stomach, but only
break down in a 7.4 pH adjusted solution of saline and media (nutrient
broth) with trypsin (a digestive enzyme) at a temperature of 370C.
These are very close to the conditions found within the definitive
host intestine. Biochemical and ultrastructural cyst studies have
shown that there is a small collagen content to the cyst wall,
but of more value is the lipid-protein complex, which would definitely
assist the passage of the metacercariae through the digestive
tract. [44, 45, 46, 47, 48, 49, 50]. A. mcintoshi cercarial penetration
glands, which assist the cercaria to enter an encystment site,
do evert for penetration in weak solutions of HCl, as shown by
Leigh [51] but otherwise do not. This indicates a mechanism which
is sensitive to gastric juices which would be encountered when
swallowed by the fish.
The question which must be asked at this point, is: "What
if these relationships, behaviors and specialized structures were
designed? " If they were designed, what would constitute
a design feature or structure for Ascocotyle or any other parasite,
and could we recognize it if we saw it? Michael Behe, in his seminal
book, [52], goes to great lengths to show this by illustrating
the bewildering complexity of the bacterial flagella, the chemical-electrical
basis for vision, and the cascade system of blood clotting. Using
the phrase "irreducibly complex", he deftly shows that
these complex systems are comprised of components, or sub-parts,
any of which, if not present, would prevent the entire system
from working, making it worthless.
He also shows the utter foolishness of expecting that a gradualistic,
Darwinian mechanism could have produced such elegant systems,
by chance, using the trial and error method, "The impotence
of Darwinian theory in accounting for the molecular basis of life
is evident not only from the analysis in this book, but also from
the complete absence in the professional scientific literature
of any detailed models by which complex biochemical systems could
have been produced...the scientific community is paralyzed. No
one at Harvard University, no one at the National Institutes of
Health, no member of the National Academy of Sciences, no Nobel
prize winner - no one at all can give a detailed account of how
the cilium, or vision, or blood clotting, or any complex biochemical
process might have developed in a Darwinian fashion."
That Michael Behe can recognize intelligent design in bacteria,
blood and vision is evident, "There is an elephant in the
roomful of scientists who are trying to explain the development
of life. The elephant is labeled 'intelligent design.' To a person
who does not feel obligated to restrict his search to unintelligent
causes, the straightforward conclusion is that many biochemical
systems were designed. They were designed not by the laws of nature,
not by chance and necessity; rather they were planned." (Emphasis
in the original). If Behe can see intelligent, planned design
in a bacterial flagella, then clearly he would see intelligent,
planned design in the HCl sensitive cercarial penetration glands
and the HCl resistant, yet trypsin and pH sensitive metacercarial
cyst of Ascocotyle which requires it to be HCl resistant at one
point in its fantastic voyage, and yet precisely trypsin and pH
sensitive at another. These and other features found within the
members of the Ascocotyle complex can be no less objects of intelligent,
planned design than Behe's bacterial flagella.
Finally, the "limitations" of hermaphrodism as understood
by an evolutionary system which would seek to amplify genetic
mixing to every extent possible does not seem to fit the ascocotylids.
Every member of this diverse group is a hermaphrodite, yet many
significant differences between species exist. Often these difficulties
are so dramatic that many new Genus and subgenus levels have been
established and discarded [53, 54, 55].
CONCLUSION
Complex obligate life cycles, as shown within the Ascocotyle complex
require that all of the special structures and features be in
place or the system will fail and the organism will not live to
bear offspring. Eggs must be ingested by a snail which will not
digest them, but rather, which will provide sanctuary for the
cercaria to develop in close proximity to the next host in the
life cycle. The second intermediate host must as well, readily
accept the infection and also be able to support a large parasite
burden with no ill effects. The cercaria must be able to penetrate
that host tissue, with a minimum of tissue response, and find
the appropriate organ for encystment. Some cercariae must have
the ability to resist gastric digestion on their way to encystment
within the intestinal lining. The metacercaria must produce an
HCl resistant, yet trypsin sensitive cyst. Finally, the hermaphroditic
adult must be able to survive in the definitive host and produce
eggs which will fall exactly where the first host may ingest them.
Chance alone cannot account for this system or the structures
it displays. An argument from intelligent design, however, might
be made on the basis of the "irreducible complexity"
of the structures and features found in this group.
ACKNOWLEDGEMENTS
I thank the reviewers for their suggestions, the late Dr. Richard
D. Lumsden for his guidance and friendship, Dr. Les Eddington
of Azusa Pacific University for free use of the EM suite, Mr.
Ronnie Palmer of the Gulf Coast Research Lab for A. pachycystis
and A. leighi specimens, Mr. Brandt Darby for help with collection
of specimens in California, Mr. Tom Keeney, Environmentalist,
Point Mugu Naval Air Weapons Station and finally, my family, especially
Patti, my wife for putting up with an 'in-home' laboratory.
FIGURES:
Figure 1. Ascocotyle mcintoshi, after Lumsden, 1963. Scale bar
= 10 microns
Figure 2. Typical redia, after Stein, 1968. Scale bar = 10 microns
Figures 3, 4. Typical cercaria, after Stein, 1968. Scale bar = 10 microns
Figure 5. Gambusia affinis, with penny for reference.
Figure 6. Fundulus parvipinnis, with penny for reference.
Figure 7. A. diminuta, SEM micrographs.
Figure 8. A. angrense, whole mount, brightfield.
Figure 9. A. leighi, encysted in fish heart, thin section.
Figure 10. A. pachycystis, encysted in fish heart, thin section.
Figure 11. A. diminuta, encysted in fish gill, thin section.
Figure 12. A. pachycystis, showing tegumental papillae.
Figure 2. Typical redia, after Stein, 1968. Scale bar = 10 microns
Figures 3, 4. Typical cercaria, after Stein, 1968. Scale bar = 10 microns
Figure 5. Gambusia affinis, with penny for reference.
Figure 6. Fundulus parvipinnis, with penny for reference.
Figure 7. A. diminuta, SEM micrographs.
Figure 8. A. angrense, whole mount, brightfield.
Figure 9. A. leighi, encysted in fish heart, thin section.
Figure 10. A. pachycystis, encysted in fish heart, thin section.
Figure 11. A. diminuta, encysted in fish gill, thin section.
Figure 12. A. pachycystis, showing tegumental papillae.
REFERENCES:
- Looss, A. Weitere Beitrage zur Kenntniss der Trematodenfauden Aegyptens. Zool. Jahrb. Syst. 12: (1899) 521-784.
- Travassos L.P. Revisao de Genero Ascocotyle Looss, 1899 (Trematoda, Heterophyidae). Mem Inst Oswaldo Cruz 23: (1930) 61-79.
- Stunkard, H.W., & Uzman, R.J. The Killifish, Fundulus heteroclitus, Second Intermediate Host of the Trematode, Ascocotyle (Phagicola) diminuta. Biological Bulletin, Marine Biological Laboratory, Woods Hole, 109: (1955) 475-483.
- Burton, P. A Review of the Trematode Genera Ascocotyle (Looss) and Phagicola (Faust) of the family Heterophyidae. Proc. Helm. Soc. Wash. 25: (1958) 117-122.
- Sogandares-Bernal, F., & Bridgman, J. F. Three Ascocotyle Complex Trematodes (Heterophyidae), encysted in fishes from Louisiana including the description of a new genus. Tulane Studies in Zoology, 8(2): (1960) 31-39.
- Sogandares-Bernal, F. & Lumsden, R. The generic status of the heterophyid trematodes of the Ascocotyle complex, including notes on the systematics and biology of the Ascocotyle angrense Travassos, 1916. J. Parasitology 49: (1963) 264-274.
- Lumsden, R.D. A New Heterophyid Trematode of the Ascocotyle Complex of Species Encysted in Poeciliid and Cyprinodont Fishes of Southeast Texas. Proc. Helminthological Soc. Wash. 30: (1963) 293-296.
- Stein, P. Studies on the life history and biology of the trematode genus Ascocotyle. Ph.D. dissertation, University of Miami, Coral Gables, Fla. (1968).
- Leigh, W.H. Observations on Life Histories of Members of the Genus Ascocotyle Looss (Heterophyidae). J. Parasitology 42:(sec2 1956) 39.
- Leigh, W.H. Life History of Ascocotyle mcintoshi Price 1936, (Trematoda Heterophyidae). J. Parasitology 60: (1974) 768-772.
- Schroeder, R. & Leigh, W. H. The life history of Ascocotyle pachycystis sp. n., a trematode from the racoon in South Florida. J. Parasitology 51: (1965) 591-599.
- Martin, W.E. Euhaplorchis californiensis, N.G., N Sp., Heterophyidae, Trematoda, with notes on its life cycle. Trans. Am. Micr. Soc. 59: (1950) 194-209.
- Burton, P. Morphology of Ascocotyle leighi, n. sp., (Heterophyidae), an avian trematode with metacercaria restricted to the conus arteriosus of the fish, Mollienesia latipinna Lesueur. J. Parasitology 42: (1956) 540-543.
- ref [11]
- Font, W., Heard, R., & Overstreet, R. Life cycle of Ascocotyle gemina n. sp., a sibling species of A. sexidigita. Trans. Am. Micr. Soc. 103: (1984) 392-407.
- ref [12]
- ref [15]
- Sogandares-Bernal, F., & Lumsden, R. The Heterophyid trematode Ascocotyle (A.) leighi Burton, 1956, from the hearts of certain poecillid and cyprinodont fishes. Zeitschr. fur. Parasitenk. 24: (1964) 3-12.
- Ostrowski de Nunez, M. Fauna de Agua Dulce en la Republica Argentina. IV. Las Cercarias de Ascocotyle (A.) tenuicollis Price, 1935, y de Pygidiopsoides pindoramensis Travassos, 1929, (Trematoda, Heterophyidae). Physis Secc. B., Buenos Aires, 35, 90: (1976) 51-57.
- Ostrowski de Nunez, M. Life history studies of heterophyid trematodes in the Neotropical region: Ascocotyle (Leighi) hadra sp. n. Mem. Inst. Oswaldo Cruz 87: (1992) 539-543.
- Ostrowski de Nunez, M. Life history studies of heterophyid trematodes in the Neotropical Region: Ascocotyle (Phagicola) diminuta (Stunkard & Haviland, 1924) and A. (P.) angrense Travassos, 1916. in Systematic Parasitology 24: (1993) 191-199, Kluwer Academic Publishers, the Netherlands.
- ref [7]
- Lumsden, R. Ultrastructure of the metacercarial cyst of Ascocotyle chandleri Lumsden 1963. Proc. Helm. Soc. Washington 135: (1968) 212-219.
- ref [18]
- ref [15]
- Leigh, W.H. The Fish Cyprinodon as the Intermediate Host for a New Species of the Trematode Genus Ascocotyle. J. Parasitology 45: (suppl 1959) 23.
- ref [10]
- ref [13]
- ref [26]
- Martin, W.E., & Steele, D. Ascocotyle sexidigita sp., n. with notes on its life cycle. Proc. Helm. Soc. Washington 37: (1970) 101-104.
- ref [10]
- Armitage, M.H. The Euryhaline Cottid Fish, Leptocottus armatus Girard 1854, Second Intermediate Host of the Trematode, Ascocotyle (Phagicola) diminuta Stunkard and Haviland 1924. Bulletin of the California Academy of Sciences 96(3): (1997) 112-116.
- Lumsden, R. Preparatory technique for electron microscopy. In: Experiments and techniques in Parasitology (eds. A MacInnis and M. Voge). Freeman Press, San Francisco pp. 215-228. (1970).
- Kobs, M. A Procedure for Preparing Microscopic Organisms for SEM. Microscopy and Analysis , Rolston Gordon Communications, Surrey, England. May 1995 issue.
- Hartl, D.L., & Clark, A.G. Principles of Population Genetics (2nd Ed.) Sinauer Associates, Inc. Publishers, Sunderland MA. 682 pp. (1989).
- McNaughton, S.J. r- and K-selection in Typha. American Naturalist 109: (1975) 251-163.
- Lights Manual: Intertidal Invertebrates of the Central California Coast, (Smith, R. and Carlton, J., eds.) University of California Press, Berkeley, CA 716 pp. (1975).
- ref [15]
- Coleman, F. Morphological and physiological consequences of parasites encysted in the bulbus arteriosis of and estuarine fish, the sheepshead minnow, Cyprinodon variegatus. J. Parasitology 79(2): (1993) 247-254.
- ref [18]
- ref [15]
- ref [30]
- ref [10]
- Lenhoff, H.M., Schroeder, R.E., & Leigh W.H. The collagen-like nature of the metacercarial cysts of a new species of Ascocotyle. J. Parasitology 46 (Suppl 1960): 36.
- ref [18]
- ref [23]
- ref [30]
- Stein, P., & Lumsden, R. An ultrastructural and cytochemical study of the metacercarial cyst development in Ascocotyle pachycystis Schroeder and Leigh, 1965. J. Parasitology 57: (1971a) 1231-1246.
- Stein, P., Lumsden, R. The ultrastructure of developing metacercarial cysts of Ascocotyle leighi Burton 1956. Proc. Helm. Soc. Washington 38: (1971b) 1-10.
- Font, W., Overstreet, R. & Heard, R. Taxonomy and biology of Phagicola nana. (Digenea: Heterophyidae). Trans. Am. Micr. Soc. 103(4): (1984) 408-422.
- ref [10]
- Behe, M.J. Darwin's black box: the biochemical challenge to evolution. The Free Press (Simon and Schuster), New York, NY. 307 pp. (1996).
- ref [2]
- ref [6]
- ref [21]