God displays His invisible attributes in creation (Romans 1:20). Therefore, creationists can use the nature of God as a guiding principle in science. One attribute of God is His desire for fellowship and community with His people (Psalm 46:10; John 3:16; Revelation 21:3-4). God's people are encouraged to show this desire. Both the Old and the New Testaments contain passages that promote friendship, fellowship, and community among believers (Exodus 12:19,47; Proverbs 17:17; 27:6,10,17; Ecclesiastes 4:12; Hebrews 10:25; Galatians 6:2; Matthew 18:15-17; Acts 4:31-32). The apostle Paul goes so far as to claim that our commitment to others should mimic that of Christ's, who laid down His life for us (Philippians 2).
If we apply the concept of "fellowship" to biology, we might expect the originally created organisms to have shown a stunningly high degree of interdependence. Because God desires His attributes to be perceived by all humans, this pattern would be expected to be prominent even after the Fall. A quick review of ecology indicates that this expectation is correct. It is possible to understand even some pathologies in this light. In a mutualistic relationship, two organisms live together and provide for the needs of the other. It is easy to see that even a slight disruption (at or after the Curse) of the mutualistic relationships might lead quickly to parasitic relationships. If we assume God created all organisms to live in mutualistic relationship (which is consistent with His value of fellowship), we could explain many modern diseases as distortions of originally beneficial relationships. As we examine disease-causing organisms, we often find organisms that appear designed to live in or on other organisms. Indeed this apparent design might be a correct perception. However it is the proper mutualistic relationship God designed and intended, not the distorted parasitic relationship brought about by the Fall.
At first glance, the blood fluke (genus Schistosoma) appears well designed for disease. To complete its lifecycle, the schistosome requires two separate hosts, with brief, free-living stages between infections. Schistosomal eggs enter a fresh water source from the urine or feces of the primary host, such as mammals or humans. These eggs hatch into ciliated larval forms called miracidia, which must infect the intermediate host, a snail, within a few hours of hatching. If a miracidium fails to find a host, it dies. After successful penetration and infection of the intermediate host (the snail), a miracidium develops into a mother sporocyst and produces a daughter generation of sporocysts. Each daughter sporocyst develops into a second larval form called a cercaria. The cercaria leaves the snail and enters the water once again, swimming until contacting the skin of the primary host. The cercaria burrows into the skin and enters blood vessels. Once in the circulatory system, it develops into the final larval form, a schistosomulum, and travels to the liver. In the liver, the schistosomulum matures into an adult schistosome and moves to a specific site of infection—blood vessels of the bladder or gut. Adult females release eggs from these sites into feces or urine for discharge from the primary host. The pathogenic effects on the human body arise when eggs lodge in blood vessels—eliciting an immune response in reaction to the foreign material, causing inflammation, ulcers, and perforation of the bowels or bladder. In very severe cases, cirrhosis or pulmonary vessel damage may occur if the eggs migrate to the liver or lungs. Symptoms of schistosomiasis include bloody diarrhea, haematuria, and dysuria.1,2
Because of the complexity of the life cycle, we infer that it arose by design. God probably created schistosomes to reproduce using this complex life cycle, or one very similar to it. The modern parasitic nature of the schistosome life cycle arose as a disruption of the fellowship between schistosome and host. If this interpretation is correct, what intermediate mechanism transformed schistosomes into parasites and what was the nature of the original relationship? Our previous study of anthrax revealed a genomic invader in the form of a pathogenic plasmid,3 while our study of mycoplasmas implicated a general decay of the organism as the generating mechanism of pathogenicity.4 Applying these principles to schistosomes, we might predict that particular details of the schistosome life cycle may show evidence of invasions, decay, or both. A few possibilities might include:
- 1. The adult schistosome might inhabit the wrong organs within the host, causing it to act differently than it would in its proper location.
- 2. The schistosome may be invading the wrong host entirely.
- 3. Modern schistosomes might produce more eggs than originally intended, causing blockages and eliciting the immune response.
- 4. The schistosome egg might have changed, eliciting an immune response, or
- 5. The host may have changed in some way, withholding some important schistosome need. In each of these possibilities, a disruption of the mutualistic relationship occurs.
To analyze these hypotheses, we need to understand the schistosome baramin, and see how modern schistosomes relate to the original blood flukes created by God and thus understand the natural variation of schistosomes. For example, if most schistosomes in the baramin do not cause disease, schistosomiasis may be a recent development of degenerating organisms. If we observe certain variations in the most virulent schistosomes, that characteristic may contribute to the cause of the disease. For example, if egg production or host specificity varied with virulence, we may reasonably conclude that these attributes contribute to the disease.
A first step in baraminology is the identification of discontinuity. The family Schistomatidae possesses a number of unique characteristics setting it apart from other flukes. Generally, flukes live in the bile duct of the liver and are hermaphroditic (each individual possesses both male and female reproductive organs). In contrast, adult schistosome worms live in blood vessels and are dioecious (having separate male and female individuals). Most flukes have a divided intestine, but the schistosome's branched intestines rejoin to form a single tube.5 Other fluke cercariae must be ingested, while the cercariae stage of the Schistosomatidae infects the primary host directly through the skin. These highly distinctive characteristics, particularly the mode of sexual reproduction, probably indicate that the blood flukes do not share an ancestor with any other worms. In ReMine's terminology,6 Schistosomatidae is probably an apobaramin.
Also, important to the study of baramins is interspecific hybridization. According to Marsh,7 the ability to produce a hybrid individual indicates that the parent species are members of the same baramin. When a person is infected with more than one species of schistosome, hybridization can occur.8 We were able to locate records of ten hybrids within the genus Schistosoma (figure 1).9,10 According to Taylor,11 hybridization within the two groups of schistosomes occurs readily,
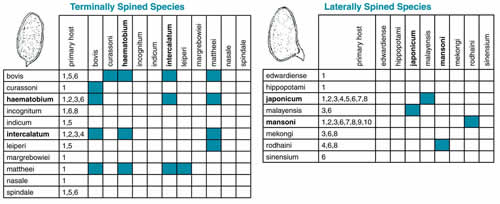
Figure 1: Schistosoma Hybridogram. Human pathogens are in bold. The numbers denote the organisms the 19 Schistosoma species inhabit. 1=Artiodactyla, 2=Human, 3=Primate, 4=Leporidae, 5=Equidae, 6=Rodentia, 7=Insectovora, 8=Carnivora, 9=Edentata, 10=Didelphis.
but the groups are largely reproductively isolated. The spines of the egg define these groups. One group has laterally arranged spines, and the other has terminally arranged spines. In ReMine's terminology, the successful hybrids suggest that these two groups are monobaramins. Future research may unite these monobaramins and clarify the identity of the baramin. Meanwhile these two monobaramins provide a basis for preliminary observations on the origin of schistosomiasis.
The variation in parasitic schistosomes gives us intriguing clues to the origin of the disease. As we might expect, all schistosomes are pathogenic to at least one animal. Human pathogens account for only four out of the 19 species in the genus Schistosoma.12 Each monobaramin contains two human pathogens and in each case the human pathogenic schistosomes can infect the greatest number of species (figure 1). This evidence is consistent with the idea that schistosomes might be invading the wrong host. Like the invading plasmids in the bacterium in B. anthracis, schistosomes might be invading systems they were not created to inhabit.
Our current research leaves us with many unanswered questions. First, the similarity between the schistosome monobaramins might constitute evidence of a single schistosome baramin, but we need further analyses to confirm this. Secondly, since all schistosomes are pathogenic, what was the original host that did not suffer from schistosomiasis? Since fellowship is a two-way street, should we examine the hosts to see if changes in anatomy or blood chemistry might be contributing to the disease? Third, we may wonder when and how fast the non-pathogenic relationship deteriorated into a parasitic one. In the case of the schistosomes, some Egyptian mummies dated to 3200 B.C. contain S. haematobium eggs.13 Since S. haematobium is considered the most recently derived group of schistosomes,14 this early date of schistosome infection indicates that the diversification of the baramin probably took place very early, perhaps even before the Flood. This study of schistosomes seems to agree with the study of anthrax in that it also seems to exhibit signs of improper invasion. It appears that "broken fellowship" has produced a nasty disease.
References
- Cox, F., ed., Modern Parasitology: A textbook of parasitology (London: Blackwell Scientific Publications, 1982), pp. 55-62.
- Rollinson, D. and Simpson, A.J.G., The Biology of Schistosomes: From Genes to Latrines (London: Academic Press, 1987), pp. 83-224.
- Wood, T.C., "Genome Decay in the Mycoplasmas," Impact 340 (2001).
- Wood, T.C., "The Terror of Anthrax in a Degrading Creation," Impact 345 (2002).
- Rollinson, ref. 4.
- ReMine, W.J., "Discontinuity Systematics: A New Methodology of Biosystematics Relevant to the Creation Model," in Proceedings of the Second International Conference on Creationism, vol. 2, edited by R.E. Walsh and C.L. Brooks (Pittsburgh, PA: Creation Science Fellowship, 1990), pp. 207-213.
- Marsh, F.L., Evolution, Creation, and Science (Washington, DC: Review and Herald Publishing Association, 1947), pp. 136-160.
- Wright, C.A., V.R. Southgate, H.B. van Wijk, and P.J. Moore, "Hybrids between Schistosoma haematobium and S. intercalatum in Cameroon," Transactions of the Royal Society of Tropical Medicine and Hygiene 68 (1974), 413-414.
- Taylor, M.G., "Hybridisation experiments on five species of African schistosomes," Journal of Helmintology 44 (1970): pp. 253-314.
- Rollinson, et al., ref. 4, pp. 362-373.
- Taylor, ref. 9.
- Boissier, J.; Desdevises, Y.; and H. Moné, "The evolution of host specificity in the Mammalia-Schistosoma host-parasite system," (submitted).
- Araújo, A. and L.F. Ferreira, "Paleoparasitology and the Antiquity of Human Host-parasite Relationships," Mem Inst Oswaldo Cruz 95 (2000): 89-93.
- Boissier, et al., ref. 12.
*The authors are at the Center for Origins Research and Education, Bryan College.




