If you have stopped receiving emails from us, please check your SPAM folder.
Also, to help stop emails from getting mistagged as spam in the future,
be sure to add 'webmaster@my.icr.org' to your contact list!
Click here for more tips!
Also, to help stop emails from getting mistagged as spam in the future,
be sure to add 'webmaster@my.icr.org' to your contact list!
Click here for more tips!

The Golden Numbers
Evolutionists theorize that the universe came into being through random means. Fundamentally, randomness lacks symmetry since the very concept of symmetry implies order. Randomness also lacks periodicity, such as events occurring in a cycle or objects in a repeating sequence. Quite simply, randomness lacks any evidence of design. Nature, however, does not display randomness.1 In fact, it exhibits quite the opposite.
In the twelfth century, Italian mathematician Leonardo Fibonacci discovered a fascinating number sequence in which each number is the sum of the previous two numbers...

APOLOGETICS
Motmot's Beauty Displays God’s Gracious Design
Beauty is God’s good gift. Whatever is truly good and beautiful comes from God (James 1:17), whose own beauty is beyond words. King David longed to know God’s magnificent beauty, saying:
One thing have...
He Doth Devise Means
“For we must needs die, and are as water spilt on the ground, which cannot be gathered up again; neither doth God respect any person: yet doth he devise means, that his banished be not expelled from him.”...




The Latest

CREATION.LIVE PODCAST
Ask, Seek, and Find with Dr. Brown | Creation.Live Podcast: Episode...
What is truth? Is truth absolute? Is it malleable as sensibilities and cultures shift?
Hosts Trey and Lauren are joined by Dr. Michael Brown to discuss...
The Golden Numbers
Evolutionists theorize that the universe came into being through random means. Fundamentally, randomness lacks symmetry since the very concept of symmetry...

Scientists Question Foundational Big Bang Assumption
In April 2024, some of the world’s leading cosmologists convened at the Royal Society in London to question the cosmological principle—the...

Moroccan Dinosaurs in Marine Rocks, Too
Two recent papers by paleontologist Nicholas Longrich and his colleagues describe some unexpected findings in phosphate mines of northern Morocco.1,2...

CREATION PODCAST
Ernst Haeckel: Evolutionary Huckster | The Creation Podcast:...
Ernst Haeckel, a German Zoologist, is famous for developing a series of images of embryos in development called Anthropogenie. These images,...

Bees Master Complex Tasks Through Social Interaction
Bees are simply incredible.1,2 These little furry fliers challenge the very foundation of Darwinism in many diverse ways.
Bees have been...
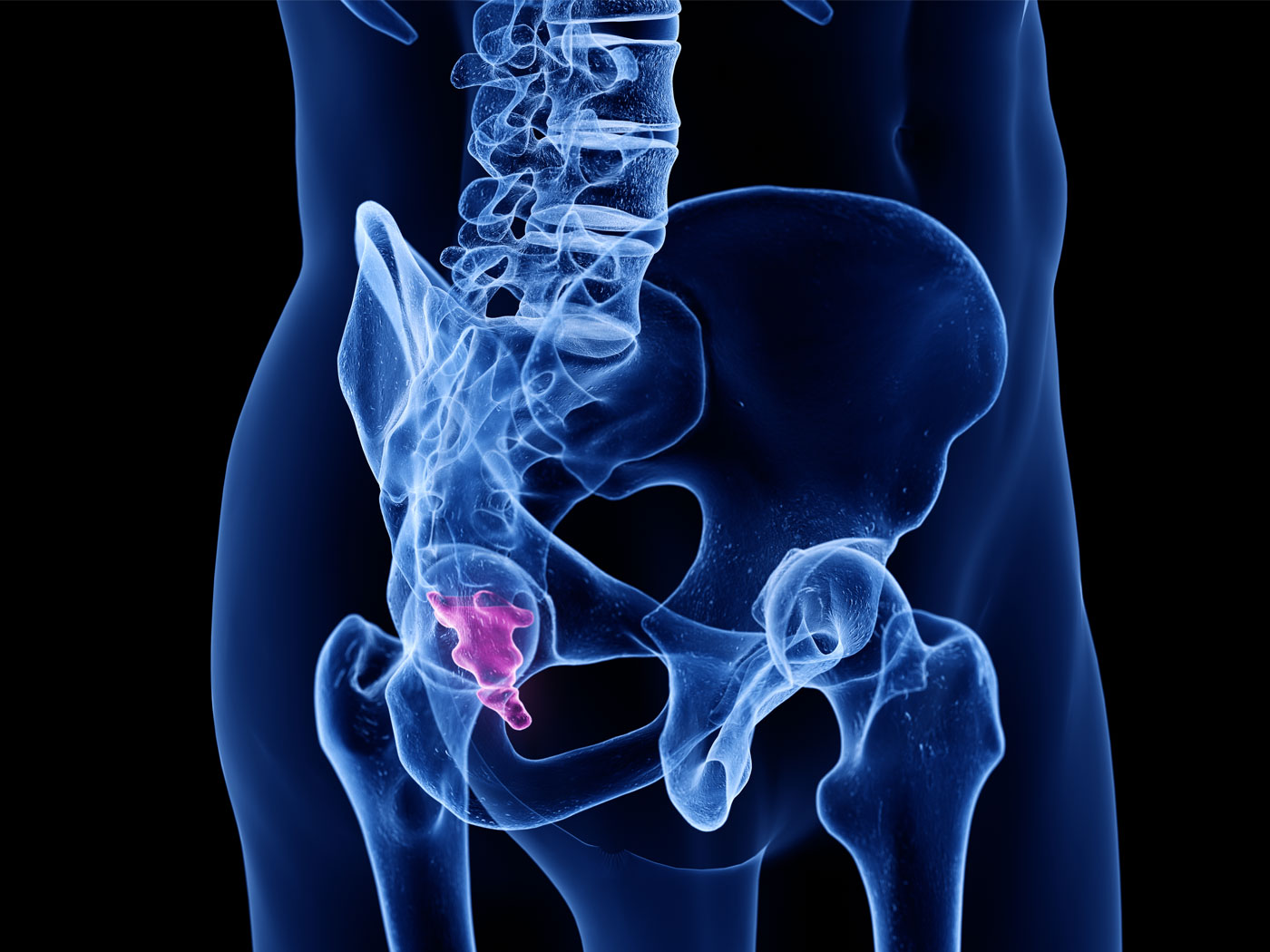
The Tail of Man’s Supposed Ancestors
Although it has been known for decades and despite insistence to the contrary from the evolutionary community, man—Homo sapiens—has never...

When Day Meets Night—A Total Success!
The skies cleared above North Texas on Monday, April 8, for a spectacular view of the 2024 Great American Solar Eclipse. Hundreds of guests joined...

The Sun and Moon—Designed for Eclipses
Before discovering thousands of planets in other solar systems, scientists tended to assume that other solar systems would be very similar to our own....

Let ICR Help You Prepare for the Great American Solar Eclipse!
On Monday, April 8th, the moon will move directly between the earth and the sun, resulting in a total solar eclipse visible in northern Mexico, much...
Creation Kids

Featured Event

Kids on Mission
Join us in the Fall of 2024! Check back soon for details of future courses.
Video
The Cost of Conviction: A Memorial Day Commemoration
These Moons Aren't Old!!! #science #creationism #podcast
ICR Dinosaur Dig Experience in Glendive, Montana!
Filling the Void #creationism #podcast
More in Video ⊳