Molluscan Methuselahs: Fossil Crassostrea Oysters
by Jake Hebert, Ph.D. | Jun. 30, 2026
 Both before and after the global Flood in the days of Noah, people routinely lived for centuries (Genesis 5 and 11). Research at ICR is finding that this extreme longevity likely characterized animals, too. For example, both fossil and modern Crassostrea oysters have been the subject of growth and longevity studies in part because Crassostrea oysters are important to the commercial seafood industry (Figure 1). And through these studies, researchers have found strong evidence of greater past longevity for these oysters. Some of this evidence has been previously discussed, but it is so remarkable that it deserves a more in-depth treatment here.1 First, I’ll address why animals living before and shortly after the Genesis Flood likely had longer lifespans. Then I’ll present the clues for greater longevity that we can expect to see in fossil remains. And finally, I’ll describe how fossil Crassostrea oysters do indeed show the expected direct and indirect evidence for very long lifespans.
Both before and after the global Flood in the days of Noah, people routinely lived for centuries (Genesis 5 and 11). Research at ICR is finding that this extreme longevity likely characterized animals, too. For example, both fossil and modern Crassostrea oysters have been the subject of growth and longevity studies in part because Crassostrea oysters are important to the commercial seafood industry (Figure 1). And through these studies, researchers have found strong evidence of greater past longevity for these oysters. Some of this evidence has been previously discussed, but it is so remarkable that it deserves a more in-depth treatment here.1 First, I’ll address why animals living before and shortly after the Genesis Flood likely had longer lifespans. Then I’ll present the clues for greater longevity that we can expect to see in fossil remains. And finally, I’ll describe how fossil Crassostrea oysters do indeed show the expected direct and indirect evidence for very long lifespans.
Before and After the Flood, Animals Likely Had Long Lives
The Bible gives no indication that the remarkable human longevity recorded in Genesis was in any way miraculous. It could have been due to biologic or environmental factors or a combination of the two. Since the first people and animals were created perfect by God, they had no genetic mistakes in their DNA. Their human and animal descendants at the time of the Flood would have had very few mistakes in their genomes. Right now, though, no one can give a definitive biological mechanism explaining the longer lifespans. Also, humans and animals were living in essentially the same pre-Flood environment, although in different ecosystems. Whether the amazing longevity was due to biologic or environmental causes or a combination of the two, it likely characterized both people and animals, including oysters.
We also know from Genesis 11 that people still had very long lifespans even after the Flood. It makes sense, then, for one to expect the same to be true of animals living shortly after the Flood.
What does this mean for people studying fossils? Nearly all the world’s fossils are the remains of creatures that were buried in mud during the Genesis Flood. But the Flood then provided ideal conditions to trigger an immediate Ice Age.2 Catastrophic Ice Age flooding, resulting from heavy rainfall and melting ice sheets, also buried and fossilized some creatures. So, one can expect both Flood and Ice Age fossils to show evidence of greater past animal longevity. There would also probably be a decline in longevity in the more recent Ice Age fossils, which are usually designated as Upper Pleistocene or Late Pleistocene by conventional scientists.3,4

Longevity, Body Size, and Maturation Times
Longevity studies in living animals show that the ones that take a long time to mature usually live longer than those that mature more quickly.5 Likewise, we know from general experience that large animals usually live longer than small animals. We see evidence of both delayed maturation and larger body sizes in fossil Crassostrea oysters. Crassostrea virginica, also known as the eastern oyster, is prized as a seafood. It is often about 5.5 inches (14 centimeters) long (Figure 1). Yet fossil Crassostrea oysters, such as C. titan and C. gigantissima (Figure 2), were twice that long.
Fossil and Modern: Same Genesis Kind?
Modern humans are the direct descendants of Noah and his family members, who had much longer lifespans than we do. Yet both we and they belong to mankind, the human “kind” or baramin (Genesis 1:20–25). In a similar way, we want to compare the lifespans of modern Crassostrea oysters to those of their direct ancestors to be able to make an accurate comparison. But not all oysters necessarily belong to the same baramin, and different oyster kinds might have different potential lifespans. Can we be reasonably sure that modern and fossil Crassostrea oysters belong to the same baramin for us to do an apples-to-apples lifespan comparison?
It complicates things that biologists and paleontologists often place very similar creatures in different taxonomic categories, even when those creatures likely belonged to the same Genesis kind. In terms of the Linnean classification system, most creationists think baramin is equivalent to either the family taxonomic level or to the level just below it, that of genus. The fact that paleontologists, despite their tendency to place similar creatures in different categories, have assigned both fossil and modern Crassostrea oysters to not just the same family but to the same genus suggests that fossil and modern Crassostrea oysters are very similar and likely really do belong to the same Genesis kind.
In fact, conventional paleontologists have explicitly stated that modern Crassostrea oysters are very likely the direct descendants of fossil Crassostrea oysters. A 1964 United States government report stated, “the [Crassostrea fossil] lineage continues, however, with little basic modification, through the Cenozoic [uppermost rocks], being represented in the Eocene by C. gigantissima (French) and probably, in modern times, by C. virginica (Gmelin).”6 The author of a 1991 paper wrote that C. gigantissima “is most certainly the direct ancestor of C. virginica.”7 Since modern and fossil Crassostrea oysters likely belong to the same baramin, it is legitimate to compare their lifespans, just as we can compare the lifespans of modern humans to those of Noah’s family.
Modern and Fossil Growth Curves
Scientists count growth bands in both living and fossil oysters and use these counts to make graphs of body size versus age. These plots show the average growth pattern for a particular oyster. These curves usually show rapid juvenile growth that gradually slows down as the creature matures. In some cases, the growth curve levels off completely, showing the age at which an oyster reaches full adult size.
It is important to consider when comparing oyster longevity, though, that bivalve lifespans can be affected by factors like water temperature and food availability.5 Bivalves living in cold water tend to live longer than bivalves living in warmer water. Limited food intake (or caloric restriction) may also increase lifespan. For this reason, when comparing fossil and modern Crassostrea lifespans, one should take care to ensure, if possible, that the fossil and modern versions lived under similar conditions, especially similar water temperatures. This again ensures an apples-to-apples comparison. In the examples described below, we can be reasonably confident that the longer lifespans of the fossil oysters were not the result of colder waters or a limited food supply.
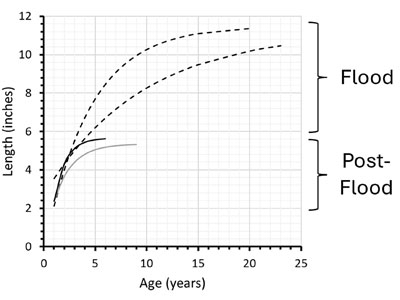
The average growth pattern and adult body size of modern C. virginica oysters from North Carolina are shown by the solid black line in Figure 3.8 Other Crassostrea studies from the United States Eastern Seaboard, as well as studies from Bangladesh and Mexico, show that modern Crassostrea lifespans of 5 to 10 years are typical, although some sources say they can live as long as 20 years in uncrowded environments.9–11
The average growth curve for Upper Pleistocene (likely Ice Age) oysters from Virginia is shown by the gray line in Figure 3.8 The Miocene (Flood) California C. titan fossil oysters (dashed black lines in Figure 3) apparently lived in pre-Flood reef environments. These California Flood oysters were twice as long, took longer to mature, and lived longer than their more recent descendants.5,8
These East and West Coast oysters came from a narrow band of about 2° latitude. At this latitude today, Pacific California coastal waters are considerably cooler than waters off the United States Eastern Seaboard. Could the great longevity of the California Miocene oysters be due to cold water? No. Conventional scientists think California coastal waters during the Miocene were considerably warmer than today. And many creationists think that pre-Flood oceans were warm compared to today’s oceans. So neither creationists nor evolutionists would likely attribute the great longevity of these California Miocene oysters to cold water.5 Likewise, the large sizes of these Miocene oysters suggest they were well-fed, thus ruling out caloric restriction as a cause for their long ages. There was a statistically significant difference between the Miocene and more recent lifespans.8
Moreover, the apparent lifespans of 20 and 23 years for these fossil Crassostrea oysters are almost certainly underestimates. Remember, these Miocene Crassostrea oysters were killed in the Flood. They might have lived even longer had the Flood not occurred. And at least one of the California Miocene fossil assemblages was “densely crowded.”8 Today, uncrowded Crassostrea oysters tend to live longer than crowded ones.9–11 So these pre-Flood oysters might have lived even longer in an uncrowded environment!
Interestingly, the maximum lifespan for Late Pleistocene (probably Ice Age) oysters (solid gray line in Figure 3) is nine years, suggesting that Crassostrea lifespans have declined since the post-Flood Ice Age, just like human lifespans shortened (Genesis 11).
More Evidence of Declining Body Size and Longevity
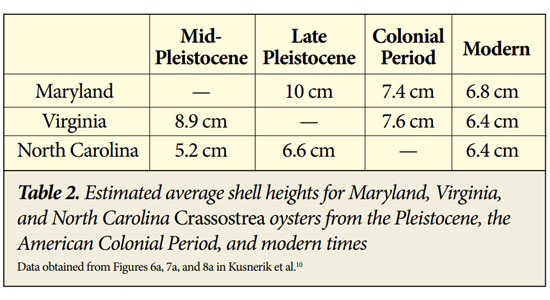
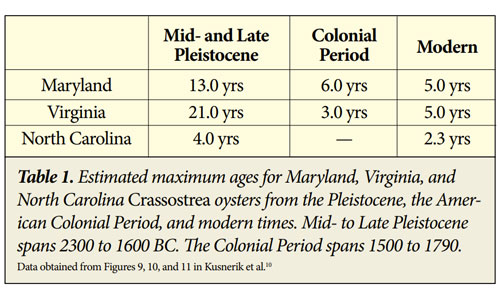
Another study of more than a thousand Crassostrea oysters from Maryland, Virginia, and North Carolina shows a general trend of decreasing longevity and shell size over time (Tables 1 and 2).10
Finally, scientists performed a study of 542 fossil and modern juvenile Crassostrea oysters from the tropical Americas. These included Pliocene and Miocene C. cahobasensis oysters, modern and Pleistocene C. columbiensis and C. virginica specimens, and an unidentified Pliocene oyster similar to C. virginica that was designated C. aff. C. virginica.
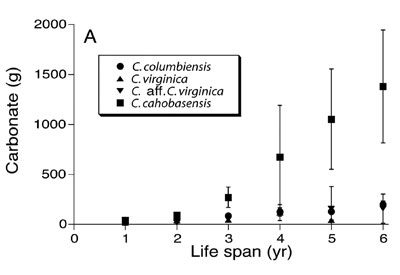
The juvenile C. cahobasensis (Miocene and Pliocene) oysters dating from the Flood were much larger and heavier at all ages than the modern and Pleistocene C. virginica and C. columbiensis juveniles (Figure 4).12 The Pliocene C. aff. C. virginica oysters might have been slightly larger than the more recent oysters, but it is difficult to tell from Figure 4. Naturally, one would expect larger juvenile body sizes to ultimately result in larger adult body sizes. So, the trend of giantism in pre-Flood Crassostrea oysters seems to have been widespread. Remember that larger animals tend to live longer than smaller animals, so this is more indirect evidence of great longevity in fossil Crassostrea oysters.
Conclusion
Fossil Crassostrea oysters lived longer, took longer to mature, and grew larger than their modern descendants. Their prolonged growth intervals are especially intriguing in light of the fact that pre-Flood humans apparently took longer than modern humans to reach sexual maturity (Genesis 5), and they probably took longer to attain skeletal maturity, as well. Thus, it makes sense that the same would have been true of pre-Flood animals.
Greater past animal longevity and delayed maturation is perfectly reasonable in light of the Bible’s claim that humans once had much longer lifespans. Who would have guessed that humble oysters could be so exciting? Bible-believing Christians should be encouraged by this. And Lord willing, we will be discussing more such evidence in future Impact articles.
References
- Hebert, J. 2024. Oysters and Pre-Flood Longevity. Acts & Facts. 53 (3): 19.
- Hebert, J. 2018. The Bible Best Explains the Ice Age. Acts & Facts. 47 (11): 10–13.
- Clarey, T. 2019. Rocks Reveal the End of the Flood. Acts & Facts. 48 (5): 9.
- Although Pleistocene fossils are usually associated with the Ice Age, in some cases, Lower or Mid-Pleistocene fossils might date from the Flood. See Holt, R. D. 1996. Evidence for a Late Cainozoic Flood/Post-Flood Boundary. Journal of Creation. 10 (1): 128–167.
- Hebert, J., R. Overman, and F. J. Sherwin. 2024. Crassostrea Oyster Fossils Show Evidence of Extreme Longevity. Creation Research Society Quarterly. 60 (3): 171–190.
- Sohl, N. F. and E. G. Kauffman. 1964. Giant Upper Cretaceous Oysters from the Gulf Coast and Caribbean. Geological Survey Professional Paper 483-H. Washington, D.C.: U. S. Government Printing Office.
- Lawrence, D. R. 1991. The Neotype of Crassostrea gigantissima (Finch, 1824). Journal of Paleontology. 65 (2): 342–343.
- Kirby, M. X. 2000. Differences in Growth Rate and Environment between Tertiary and Quaternary Crassostrea Oysters. Paleobiology. 27 (1): 84–103.
- Harzhauser, M. et al. 2016. Age Structure, Carbonate Production and Shell Loss Rate in an Early Miocene Reef of the Giant Oyster Crassostrea gryphoides. Biogeosciences. 13: 1223–1235.
- Kusnerik, K. M. et al. 2018. Using the Fossil Record to Establish a Baseline and Recommendations for Oyster Mitigation in the Mid-Atlantic U.S. In Marine Conservation Paleobiology, Topics in Geobiology, volume 47. C. L. Tyler and C. L. Schneider, ed. New York, NY: Springer.
- Osborne, P. Crassostrea virginica. Animal Diversity Web. Posted on animaldiversity.org, accessed April 10, 2026.
- Kirby, M. X. and J. B. C. Jackson. 2004. Extinction of a Fast-Growing Oyster and Changing Ocean Circulation in Pliocene Tropical America. Geology. 32 (12): 1025–1028.
* Dr. Hebert is a research scientist at the Institute for Creation Research and earned his Ph.D. in physics from the University of Texas at Dallas.
Stage image credit: Susan Windsor
Adaptive Trait Variation Conferred by Engineered Genetic Diversity
 Global environments are highly diverse and dynamic, offering many changes and adaptive challenges to creatures. However, DNA sequence variability engineered into plant and animal genomes is one design feature (of many) that allows creatures to rapidly deploy adaptive traits in response to sensing a broad range of environmental conditions.
Global environments are highly diverse and dynamic, offering many changes and adaptive challenges to creatures. However, DNA sequence variability engineered into plant and animal genomes is one design feature (of many) that allows creatures to rapidly deploy adaptive traits in response to sensing a broad range of environmental conditions.
An Introduction to Genetic Diversification
More...Engineered Genomic Changes in Adaptation
 Programmed genome rearrangements (PGRs) are deliberate, genetically controlled changes in an organism’s DNA sequence and chromosome structure that occur during normal development or in response to detecting changes in environmental conditions. These PGRs range from single bases to genes to even large genomic regions. They are precisely orchestrated modifications that involve deletions, inversions, duplications, translocations, sequence reductions, or ...More...
Programmed genome rearrangements (PGRs) are deliberate, genetically controlled changes in an organism’s DNA sequence and chromosome structure that occur during normal development or in response to detecting changes in environmental conditions. These PGRs range from single bases to genes to even large genomic regions. They are precisely orchestrated modifications that involve deletions, inversions, duplications, translocations, sequence reductions, or ...More...
Novel Orphan Genes Aid in Regulated Adaptation
 Orphan genes (OGs) are genes that are unique to a specific kind of creature. This is especially significant when creatures that are considered evolutionary ancestors lack these genes. In other words, OGs have no discernable evolutionary ancestry but appear suddenly without any evolutionary precursors—debunking the story of gene evolution and biological evolution in general. And even more interesting is that many OGs play significant roles in environmental ...More...
Orphan genes (OGs) are genes that are unique to a specific kind of creature. This is especially significant when creatures that are considered evolutionary ancestors lack these genes. In other words, OGs have no discernable evolutionary ancestry but appear suddenly without any evolutionary precursors—debunking the story of gene evolution and biological evolution in general. And even more interesting is that many OGs play significant roles in environmental ...More...
Pseudogenes Are Not Pseudo Anymore
Introduction
One of the past arguments for evidence of biological evolution in the genome has been the concept of pseudogenes. These DNA sequences were once thought to be the defunct remnants of genes, representing nothing but mutated genomic “fossils” in the genomes of plants, animals, and humans. However, much research has been published over the past several decades showing that pseudogenes are not only functional but key to organism survival.1,2
More...More Articles
- Long Non-Coding RNAs: The Unsung Heroes of the Genome
- Genomic Tandem Repeats: Where Repetition Is Purposely Adaptive
- The 3-D Genome: A Marvel of Adaptive Engineering
- Gene Complexity Showcases Engineered Versatility
- RNA Hoops: When Circular Reasoning Makes Sense
- Engineered Parallel Gene Codes Defy Evolution
- Genetic Recombination: A Regulated and Designed Chromosomal System
- Galápagos Finches: A Case Study in Evolution or Adaptive Engineering?
- RNA Editing: Adaptive Genome Modification on the Fly
- Small Heritable RNAs Pack a Big Adaptive Punch
- Trait Variation: Engineered Alleles, Yes! Random Mutations, No!
- Transposable Elements: Genomic Parasites or Engineered Design?
- Epigenetic Mechanisms: Adaptive Master Regulators of the Genome
- Jupiter's Young Moons