If you have stopped receiving emails from us, please check your SPAM folder.
Also, to help stop emails from getting mistagged as spam in the future,
be sure to add 'webmaster@my.icr.org' to your contact list!
Click here for more tips!
Also, to help stop emails from getting mistagged as spam in the future,
be sure to add 'webmaster@my.icr.org' to your contact list!
Click here for more tips!

Scientists Question Foundational Big Bang Assumption
In April 2024, some of the world’s leading cosmologists convened at the Royal Society in London to question the cosmological principle—the assumption that the universe is the same everywhere and in every direction.1,2 This is a highly significant development since the cosmological principle is a foundational assumption of the Big Bang model for the universe’s origin.
The cosmological principle assumes that the universe is both homogenous and isotropic. Homogeneity means the distribution of matter and energy in the universe is roughly uniform if one looks at the...

Dolomite Problem Best Solved by Flood
Dolomite is a very common sedimentary rock, comprising about 30% of all carbonate rocks.1,2 Its chemical formula is MgCa(CO3)2, whereas the more common limestone is CaCO3. Oddly, ocean water is supersaturated...
Fruitless Trees
“Woe unto them!...trees whose fruit withereth, without fruit, twice dead, plucked up by the roots.” (Jude 1:11-12)
Many illustrations in Scripture compare the responsibility of trees to bear fruit and...




The Latest

The Golden Numbers
Evolutionists theorize that the universe came into being through random means. Fundamentally, randomness lacks symmetry since the very concept of symmetry...
Scientists Question Foundational Big Bang Assumption
In April 2024, some of the world’s leading cosmologists convened at the Royal Society in London to question the cosmological principle—the...

Moroccan Dinosaurs in Marine Rocks, Too
Two recent papers by paleontologist Nicholas Longrich and his colleagues describe some unexpected findings in phosphate mines of northern Morocco.1,2...

CREATION PODCAST
Ernst Haeckel: Evolutionary Huckster | The Creation Podcast:...
Ernst Haeckel, a German Zoologist, is famous for developing a series of images of embryos in development called Anthropogenie. These images,...

Bees Master Complex Tasks Through Social Interaction
Bees are simply incredible.1,2 These little furry fliers challenge the very foundation of Darwinism in many diverse ways.
Bees have been...
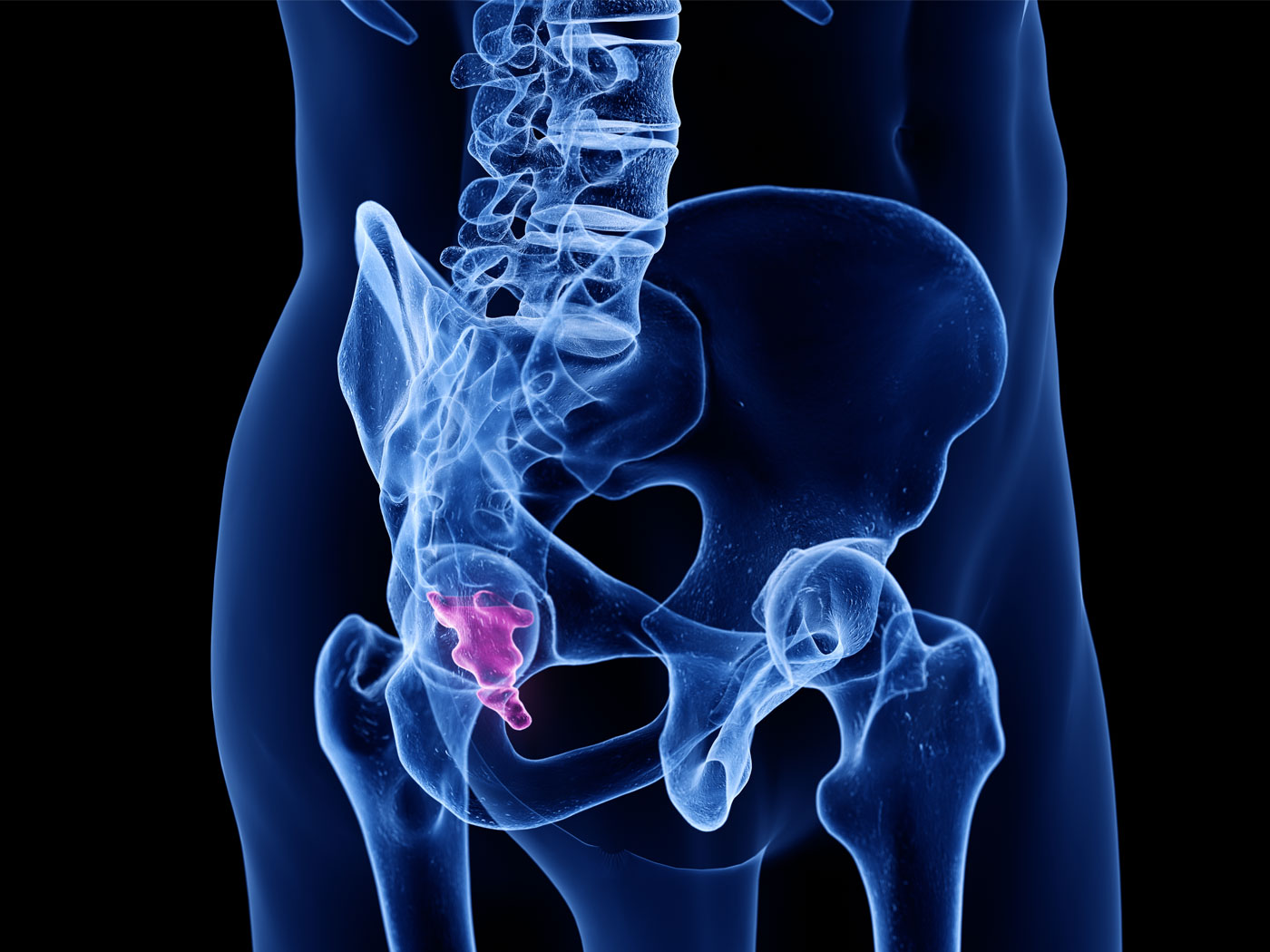
The Tail of Man’s Supposed Ancestors
Although it has been known for decades and despite insistence to the contrary from the evolutionary community, man—Homo sapiens—has never...

When Day Meets Night—A Total Success!
The skies cleared above North Texas on Monday, April 8, for a spectacular view of the 2024 Great American Solar Eclipse. Hundreds of guests joined...

The Sun and Moon—Designed for Eclipses
Before discovering thousands of planets in other solar systems, scientists tended to assume that other solar systems would be very similar to our own....

Let ICR Help You Prepare for the Great American Solar Eclipse!
On Monday, April 8th, the moon will move directly between the earth and the sun, resulting in a total solar eclipse visible in northern Mexico, much...

Total Eclipse on April 8th
“You alone are the LORD; You have made heaven, the heaven of heavens, with all their host, the earth and everything on it, the seas and all that...
Creation Kids

Featured Event

Kids on Mission
Join us in the Fall of 2024! Check back soon for details of future courses.
Video
ICR Dinosaur Dig Experience in Glendive, Montana!
Filling the Void #creationism #podcast
We Are More Than Animals! #creationism #podcast
Ernst Haeckel: Evolutionary Huckster
More in Video ⊳